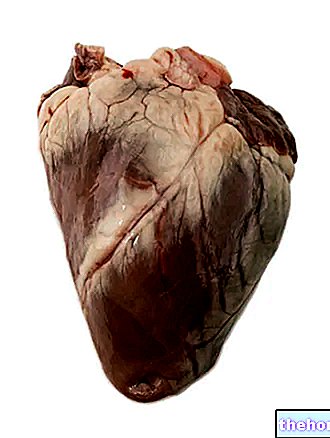
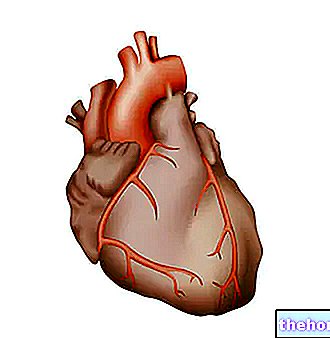
Cykl Krebsa jest również nazywany cyklem kwasów trikarboksylowych i wykorzystuje acetylokoezim A jako wyjściowy metabolit, który jest otrzymywany w wyniku działania dehydrogenazy pirogronianowej na pirogronian wytwarzany przez glikolizę.
ATP i moc redukującą uzyskuje się z cyklu Krebsa; siła redukująca jest przesyłana do łańcucha oddechowego, gdzie NADH i FADH2 są utleniane odpowiednio do NAD+ i FAD: siła redukująca jest przekazywana wzdłuż łańcucha oddechowego do układów sprzęgających, z których wytwarzany jest dalszy ATP.
Cykl Krebsa jest ośrodkiem nerwowym nie tylko metabolizmu glukozy, ale także kwasów tłuszczowych i aminokwasów, w rzeczywistości pirogronian, który jest przekształcany w acetylokoenzym A, nie pochodzi tylko z degradacji glukozy: jest otrzymywany m.in. na przykład, również z transaminacji alaniny (aminokwasu).
Około 80% acetylokoenzymu A, który bierze udział w cyklu Krebsa, pochodzi z metabolizmu kwasów tłuszczowych.
Koenzym acetylowy A jest tioestrem, dzięki czemu ma wysoką zawartość energii, która jest wykorzystywana przez syntaza cytrynianowa aby utworzyć nowe wiązanie węgiel-węgiel; syntaza cytrynianowa jest pierwszym enzymem cyklu Krebsa.
Węgiel metylowy acetylokoenzymu A chętnie uwalnia (poprzez tautomerię) proton (staje się karboanionem) i atakuje węgiel karbonylowy szczawiooctanu: powstaje wysokoenergetyczny tioester (koenzym cytrylowy A), z którego w wyniku hydrolizy otrzymuje się cytrynian i koenzym A. Syntaza cytrynianowa jest modulowana ujemnie przez produkt, tj. przez cytrynian i przez ATP: jeśli cytrynian się akumuluje, oznacza to, że ten etap jest szybszy niż inne, dlatego musi spowolnić (cytrynian jest negatywny modulator).

ATP wpływa również na działanie syntazy cytrynianowej, ponieważ siła redukująca jest uzyskiwana z cyklu Krebsa, który jest następnie przesyłany do łańcucha oddechowego, z którego wytwarzany jest ATP; jeśli ATP się kumuluje, oznacza to, że powstaje więcej niż potrzeba. Spowalniając cykl Krebsa (cykl spowalnia, jeśli jeden z jego etapów jest spowolniony) produkcja ATP jest również spowolniona: ujemna modulacja ATP jest modulacją sprzężenia zwrotnego (powstawanie jednego z produktów końcowych jest modulowana poprzez dostosowanie szybkości jednego kroku w procesie).
W drugim etapie cyklu Krebsa cytrynian jest przekształcany w izocytrynian pod wpływem enzymu akonitaza; nazwa enzymu wywodzi się z faktu, że cytrynian jest najpierw odwadniany z wytworzeniem cis-akonitynianu, a następnie woda ponownie wchodzi do wody, przyłączając się do węgla innego niż ten, z którym była wcześniej związana. Izocytrynian uzyskuje się bez opuszczania przez substrat miejsca katalitycznego; akonitaza jest enzymem stereospecyficznym: rozpoznaje trzy centra karboksylowe cytrynianu, co powoduje, że cytrynian pozostaje związany z enzymem, tak że wyjście i wejście wody zawsze mija przez półprodukt cis-akonitianowy.
W trzecim etapie cyklu Krebsa pojawia się pierwszy czynnik energetyczny, ponieważ następuje utrata węgla eliminowanego jako dwutlenek węgla. Enzymem, który katalizuje ten etap jest dehydrogenaza izocytrynianowa; substrat ulega przede wszystkim odwodornieniu: NAD+ uzyskuje moc redukującą i powstaje szczawianobursztynian (jest to szczawiowa pochodna kwasu bursztynowego), który następnie ulega dekarboksylacji do α-ketoglutaranu.
Enzym dehydrogenaza izocytrynianowa ma dwa miejsca modulacji: dodatnią modulację z powodu ADP i ujemną modulację z powodu ATP.Ilość spożywanego dziennie ATP jest bardzo wysoka: ATP dostarcza energię uwalnianą przez jego hydrolizę, „ADP i cały” ortofosforan.
Całkowite stężenie nukleozydów (zasada azotowa plus cukier) i nukleotydów (nuklozyd plus fosforan) w organizmie jest prawie stałe: powiedzieć zatem, że c "to dużo ATP lub mało ADP (lub odwrotnie, dużo ADP i małe ATP) to to samo, ADP jest synonimem zapotrzebowania na energię, a zatem jest modulatorem dodatnim, natomiast ATP jest objawem dostępności energii, a zatem jest modulatorem ujemnym.
KONTYNUUJ: Część druga”